

I'm Going To Leave This Post Here For A While (Until I Find A Place For It). This Is The One On The Y-chromosome And Mitochondrial Lineage Of Polynesia. Like Usual, It's A Lot To Read And A Little Time Consuming (And Straining On The Eyes), But Your Time, Energy, And Effort Are Worth It Because It's Interesting And Accurate. ENJOY.










I sat in Lucy's cafe in Avarua, the capital (indeed, the only town) of Rarotonga, having an ice-cream, just looking at the people coming and going. Did they look more Asian or more American? It wasn't obvious to me. I distinctly remember one small girl who could have come straight from a National Geographic cover story on the Amazonian rainforests. If only I could test the mitochondrial DNA of the people in the cafe! I was sure I'd be able to tell whether their closest genetic links were with Asia or America. So, at the next hospital appointment to review my fracture shoulder, I explained that I was a geneticist and what I had in mind. Somehow or other I managed to persuade the hospital to let me have the remnants of thirty-five blood samples left over from blood-sugar tests. Diabetes is very common in Rarotonga, and so there are a lot of tests for blood glucose levels. I stored the samples in the freezer back at the hotel. After my shoulder healed - A little too quickly, I thought - I carried the precious phials of blood with me to Australia, where I very nearly had them confiscated by customs, then eventually back to England and my laboratory.
The day after I got back, I unwrapped the samples. There was blood oozing everywhere. The glass tubes had smashed - but fortunately, not all of them. Twenty were still intact, and I got on with sequencing their mitochondrial DNA. Nowadays, DNA sequencing is done automatically in extremely expensive machines, but in the early 1990s it was a manual operation which involved tagging fragments of DNA with mildly radioactive isotopes and separating them in an electric field. There was a point at the end of the long process when the X-ray film which recorded the pattern of radioactive bands that revealed the sequence slowly issued from the developing machine. It was always a tense moment. Will there be a good set of bands? Will there be any bands at all? If the bands are too weak or absent altogether, then something has gone wrong and it's back to the laboratory bench for another three days.
This time, with the first ten of the twenty samples, everything had worked. Drawn across the X-ray film were four wide columns of dark bands, like bar codes, where the weak radioactivity had blackened the photographic emulsion. The four columns were each divided into ten tracks, one for each sample. Each of the four columns read the sequence of one base, so by putting them together the complete sequence could be worked out. I had arranged it this way, with the ten samples side by side, so that it was easy to see where the differences between individuals were. That was what I really wanted to focus on - the differences between people, rather than the similarities. A straight line across all ten tracks means that all ten samples were identical at that position; a line with interruptions meant that some samples were different at that position.
In the lab we had sequenced ourselves and a few friends, mostly European, and typically there would be a couple of dozen lines in each batch of ten samples that showed these tell-tale interruptions. When the Rarotongan film came sliding out of the developer there were bands all right, but there was not a single interruption. They were all exactly the same. Had I made a mistake? Had I inadvertently combined the samples somewhere along the line? I needed to develop the second film from samples 11-20 to find out. When this came out it looked at first as if I really had made a mistake. Another whole film of straight lines. But then I saw one track, one individual, that was different from all the rest. Very different. And three other tracks had a single interrupted line. So they hadn't been mixed. They were real results. I realized at once that they were stunning, and that before very long I would have the answer to the origin of the Polynesians.
Reading through the sequences more carefully and comparing them with the European reference sequence, I saw that the major sequence shared by sixteen of the twenty Polynesians was different at four positions: 189, 217, 247, and 261. The similar sequence shared by three individuals on the second film differed from this only in that they did not have the variant at 247. Otherwise their mitochondrial DNA was identical; they had to be very closely related to the first sixteen. But the twentieth sample was completely different. It had nine variants from the reference sequence along the control region, none of which was shared with the main Rarotongan cluster. Since the blood samples had come from the outpatient clinic in Avarua, there was no guarantee that they were from native Rarotongans, and so I assumed that this unusual sequence was from a tourist or a visitor from another part of the world. Since very few mitochondrial sequences had been published in 1991, there was no telling where on the globe this might be.
I concentrated on the main result - the astonishing similarity of nineteen of the twenty samples. This had to be the mitochondrial DNA of the original Polynesians. All we had to do to solve the Polynesian question was to look in both south-east Asia and South America for comparisons. If we found DNA matches in Chile of Peru, or even coastal North America, then Heyerdahl was right. If we found them in south-east Asia, he was wrong. If we didn't find a match in either region, then everyone was wrong. Whichever turned out to be true, one thing was certain: we were going to settle, once and for all, the debate that had raged for over two hundred years. I started to plan my next trip.
You might be asking yourself at this point: 'Surely if it was as easy as that, blood groups would have given the answer long ago?' It isn't as if blood groups of Polynesia had never been studied; indeed, the first results from Samoa in central Polynesia had been published in 1924, only five years after the Lancet paper by the Herschfelds which first introduced the potential of blood grouping in anthropology. The south Pacific, as I was fast learning, had been a popular place for scientific fieldwork for a long time. However, while they formed a plank in the argument in favour of a south-east Asia origin, decades of work on blood groups and other classical genetic systems had still not produced a definite answer to the puzzle, first because the variations are not definitive, and second because the evolutionary relationships between the types are not known. For example, Polynesians, native South Americans and south-east Asians all have a high frequency of blood group O. Polynesians also have quite a high frequency of blood group A, which is virtually absent in South America. But they also have a low frequency of blood group B, which is quite common in south-east Asia. So what can you make of all that? Which theory do these data support? Advocates of the Asian origin would argue that the extreme rarity of blood group A in native South Americans means that the Polynesian blood group A couldn't have come from South America. Supporters of the South American case could legitimately respond by saying, as Arthur Mourant suggested in 1976, that the Polynesian blood group A came originally not from Asia but from Europeans through intermarriage over the last three hundred years. And anyway, where's all the blood group B that should have come from Asia? Add to all this uncertainty the fact that, ultimately, all native Americans trace their origins to Asia through the settlers who crossed the Bering land bridge thousands of years before, and you have a complete mess. Blood group O could have reached Polynesia either directly from Asia of via the Americas. There is no way of knowing. With only three blood group genes - A,B, and O - certainty remains out of reach.
Other classical genetic markers are more variable, none more so than the one that controls the tissue type system important in organ transplantation. Just as blood needs to be cross-matched before a transfusion to avoid a fatal immune reaction, so you must match tissue types between donor and recipient when transplanting organs like heart, kidneys or bone marrow. You don't hear of people waiting for a blood transfusion because they can't find a match, but it is a a sadly familiar story to hear of patients waiting for months or even years for a suitable heart or kidney donor, often dying before one is found. This is because while there are only four blood groups (A, B, AB, and O) there are scores of different tissue types.

I must admit here and now to a serious personal weakness. I have a complete mental block when confronted with the bewildering variety of tissue types. Some of my best friends are cellular immunologists who live, work and breathe tissue types, and the Institute where I work is packed with them. Yet something switches off in my brain when they start describing the various types. All of them begin with the three letters HLA. Then numbers and letters are tacked on to the end: HLA -DRB1, HLA-DPB2, HLA-B27 and so forth. Time and again I go to seminars which kick off with a slide showing a table of this horrendous alphanumeric melange. For years I concentrated, thinking it would sink in eventually if I tried hard. After all, have to teach this stuff in my genetics classes. But to no avail. I reluctantly conclude that I am genetically incapable of understanding tissue types beyond knowing that there are an awful lot of them.Which, fortunately, is all you need to know as well. Since there are lots them, and there are quite a lot of data from Polynesia, South America, and south-east Asia, it is relatively easy to track them; and sure enough, most of the tissue type connections are between Polynesia and Asia. But not all. A type called HLA-Bw48 is very rare except among Polynesians, Inuit and native North Americans. However, though there is certainly plenty of variation, the evolutionary connection between the different types was not known. So, for example, you couldn't tell whether HLABw48, the type found also in North America, was closely related to other Polynesian types or not. Compare that to the situation with the mitochondrial DNA from Rarotonga. We know that there are three types; we also know that two of them are very closely related to each other, while the third is not. That, as we will see, is an enormous help. We can search other lands not only for the Polynesian mitochondrial types themselves, but for others that are closely related to them as well.
By the time I had planned the return trip, and persuaded the Royal Society to pat for it - after all, they had paid for Cook's first voyage to Tahiti, as I pointed out in my application - data from native North and South Americans produced by other researchers had begun to circulate. Just as there was one cluster in the Rraotongan sample (if we include the two closely related types in a single cluster and forget about the single sequence from the 'tourist'), so there were four main clusters in the Americas. Three of these had quite different mitochondrial DNA sequences; the fourth was rather similar to the main Rarotongan sequence of 189, 217, 247, 261, but with variants at positions 189 and 217 only. This looked very interesting. Moreover, both the native American and Rarotongan DNAs shared another unique feature. At the opposite side of the mitochondrial DNA circle from the control region that we had sequenced, a small piece of DNA, only nine bases long, was missing. This definitely increased the chances that the American and Polynesian types were related. Things were looking up for Heyerdahl.
I heard that in Hawaii Rebecca Cann, one of the authors with Allan Wilson of the origial 1987 paper on mitochondrial DNA and human evolution, was studying the DNA of native Hawaiians. This is hard work because, unlike in Rarotonga, there are very few of them left. Two hundred years of immigration, mainly from Asia and America, have reduced the native Hawaiians to a fringe population, many of them living a marginal existence - an all too familiar legacy of colonialism. However, schemes have recently been introduced by which special grants and scholarships are awarded to those who can prove they are of native Hawaiian ancestry. One way of proving this ancestry is through DNA testing; so there was an extra incentive to find out about the mitochondrial genetics of the native Hawaiians.
On my return visit to Rarotonga I arranged to call on Becky Cann in Hawaii, where we sat down in her lab with her postgraduate student, Koji Lum, to compare results. It didn't take long to discover that we had both found the same major Polynesian type, with the deletion and the same control region variants. This was very exciting, and confirmed the link between the people of Hawaii and those of Rarotonga, three thousand miles to the south. Already I was imagining the enormous ocean distance that separated the two island groups, and the fantastic voyages that must have carried these genes across the sea. Even though it was not unexpected, given the wealth of the evidence from the days of Captain Cook onwards that connected all the Polynesians to a common ancestry, just seeing the proof was thrilling. Reluctantly, Becky left to prepare for a seminar, leaving Koji and me in the office sharing our admiration for the voyages of the Polynesians that had carried these genes to Rarotonga and Hawaii.
What followed is one of those rare moments in science when something is revealed that has never been seen before. I was about to pack away my data when I remembered the unusual Rarotongan sequence that I had interpreted as belonging to a tourist and more or less forgotten about. I returned to Koji and asked him if he had ever seen anything like it in native Hawaiians. He agreed to have a look and unpacked his own sheets of results. There was one that stood out from the rest. I laid out my sheet, rather like a roll of wallpaper - this was in the days before laptops - on which the Rarotongan sequences were drawn out, and soon located the unusual sequence. At first Koji's and my sequences looked completely different; then we realized that we had been reading them from opposite ends. I turned mine around, and began to go through the strange Rarotongan sequence. I read from the left-hand side. The first variant was at position 144.
'Do you have anything with 144?' I asked.They were identical. We both looked up at the same time. Our eyes met and two huge, silent smiles shone out from our faces. This was not the DNA of a tourist at all. Discounting the remote possibility that I had accidentally collected a blood sample from a native Hawaiian on holiday in Rarotonga, this had to be a second genuine Polynesian DNA type that had reached into the Pacific as far as the Cook Islands and Hawaii. But where had it come from? It would take another six months to find out.
'Yes' said Koji.
I carried on four more bases to 148. 'Anything at 148?'
'Yes, in the same sample,' he replied.
I could feel the thrill of discovery tingling up my spine. I carried on. '223?'
'Yes.'
'241?'
'Yes.'
I accelerated. '293?'
'Yes.'
'362?'
'Yes.'
I flew down to Rarotonga, more determined than even that we would solve the mystery surrounding the origins of the Polynesians. When I got there, Malcolm, my host from my first visit arranged for me to meet the man who ran the Prime Minister's office. In most countries this would be quite impossible, but in Rarotonga it was accomplished at Malcolm's Christmas party on the beach. It was fortunate that I met Tere Tangiiti and arranged an appointment early on in the proceedings; because my abiding memory of that party was not of making a crucial diplomatic contact, but of the colour blue: the color of Curacao which, mixed with champagne, makes the cocktail Blue Lagoon. Blue Lagoon, seafood omelettes and my digestion don't mix. I was soon to discover the interesting scientific fact that whatever it is they use to colour Curacao, it is not destroyed in the human stomach. Ten years later I steel feel sick at the sight of it.
I needed to get the permission of the cabinet and the cooperation of George Koteka at the health department to collect a substantial DNA sample from Rarotonga and the other islands. I met the cabinet in the Prime Minister's office above the post office, and they could not have been more helpful. Within a few weeks I had collected five hundred samples from Rarotonga, Atiu, Aitutaki, Mangaia, Pukapuka, Rakahangha, Manihiki and even from the tiny atoll of Palmerston (population sixty-six). I packed them carefully in ice and took them back to Oxford.
The institute of Molecular Medicine, where my laboratory is based, is built around the pioneering work of its first director, Professor Sir David Weatherall. His research over the past twenty-five years has been focused on the inherited diseases of the blood, in particular those involving the main component of red blood cells - hemoglobin. These diseases are not particularly common in northern latitudes, but have a quite devastating effect on public health in parts of Africa, Asia, and Mediterranean Europe. The main diseases, sick cell anaemia in Africa south of the Sahara and thalassaemia in Asia and Europe, kill hundreds of thousands of children every year. The causes of all this misery are tiny mutations in the haemoglobin genes, which very slightly alter the oxygen-carrying properties of the red blood cells. In sickle cell anaemia, the usually circular red blood cells visually change shape, as the name implies, and can no longer slide past each other in the narrowest of blood vessels. This clogs up the flow of blood to vital tissues. In thalassaemia the haemoglobin itself forms clumps inside the red blood cells, which are often destroyed in the spleen. Either way the anaemias can be fatal if left untreated; and still the only effective remedy is repeated blood transfusions which - quite apart from the side-effects they cause by overloading the body with iron - are beyond the public health budgets in most of the affected regions.
Why do these diseases occur in some places and not in others? The answer is - malaria. Sickle cell anaemia and thalassaemia are found principally in parts of the world where malaria is, or has been, endemic. Both diseases, in order to develop, require a double dose of the mutant haemoglobin gene, one from each parent. Many inherited diseases follow the same pattern; among Europeans the most familiar is cystic fibrosis, where the parents are carriers with one copy each of the mutant gene but no symptoms of the disease. For a reason that even now is not entirely clear, the parasite that causes malaria finds it difficult to infect the red blood cells of sickle cell anaemia and thalassaemia carriers, who as a consequence become at least partially resistant to the disease. Over many generations, this resistance leads to a spread of the haemoglobin mutations in the malarial regions through the forces of natural selection. However, while the mutations are good for carriers, they can be devastating for their children, because some of the offspring of two carrier parents get the double dose of haemoglobin mutants and develop the potentially fatal anaemias. This cruel balance of carrier advantage and offspring elimination keeps the haemoglobin mutants at a high frequency wherever malaria is found. Malaria does not cause these diseases directly, but does do indirectly by allowing, indeed, encouraging, the haemoglobin mutations - which are the real cause - to survive and prosper. So, even if you eliminate malaria, you do not at once eliminate these diseases. In Mediterranean Europe - Sardinia, Italy, Greece, Cyprus, and Turkey - programmes to eradicate the mosquitoes which carry the malarial parasite have virtually eliminated malaria - but not thalassaemia. Tens of thousands of people still carry the haemoglobin mutations, and only an entirely different programme, built around the genetic testing of prospective parents to see if they are carriers, is reducing the incidence of the disease.
Many people from the Mediterranean have emigrated to different parts of the world, in particular to the United States and Canada, Australia and Britain. With them, literally inside them, travel the thalassaemia genes, so that the disease is also encountered in these communities. For the same reason, forced immigration on slave ships from West Africa introduced the sickle cell gene to North America, where sickle cell anaemia is still encountered, even though there is no malaria. Gradually, over many generations, it will fade from these populations as the mutations are eliminated either by active counseling programmes or simply by the deaths of those who have the disease. Without malaria to help it along it will suffer the ultimate fate of all disease genes - extinction by natural selection.
Unravelling the roots of sickle sell anaemia and thalassaemia has had a major influence on genetics. It is no exaggeration to say that without the examples of these two diseases to guide reseachers, very few of the great advances that have been made since the mid-1980s in finding the causes of genetic diseases would have happened. It was studies of the inherited anaemias that convinced scientists and doctors that simple mutations in genes did indeed cause disease.
The advantages of all this work for me, in my search for the origins of the Polynesians, were far more prosaic. It was field work in the islands of south-east Asia and Oceania, mainly Papua New Guinea, Vanuatu, and Indonesia, that finally proved the connection between thalassaemia and malaria. The thalassaemia genes were found only in the low-lying, swampy areas near the coast, where malaria was common, while in the mountainous interiors, where the mosquitoes could not surive the high aliude, the troublesome genes were virtually absent. As a result of this research, the freezers in the Institue of Molecular Medicine were full of DNA samples from these islands. I needed to look no further than the first floor of the Institute where I worked to augment my own samples from Polynesia with a fabulous collection which covered more or less the entire route from south-east Asia into the remote Pacific. If the Polynesians had come that way, surely we would find their mitochondrial DNA scattered along the route.
Over the summer of 1992 I sequenced over 1,2000 mitochondrial DNAs. The first thing to do was to see whether we could find any with the small deletion. Nineteen out of the twenty Rarotongans were missing this tiny segment and it was very easy to test for it. And there it was: very common in Samoa and Tonga; less common further west in Vanuatu and the coast of New Guinea. The deletion was even less frequent in Borneo and the Philippines, but still there far to the west among the native Taiwanese. This looked like good evidence for an Asian origin. But remember that we knew from published work that the same tiny deletion was also to be found in North and South America. Were we going to find ourselves in the same frustrating situation as everyone else who had tried to use genetics to solve the puzzle, unable to differentiate between a gene that had arrived in Polynesia directly from Asia and one imported indirectly via the land bridge to America? Our only hope was that the control region sequence itself would be able to tell the difference.
The common sequence in Rarotonga, and from the lab in Hawaii, had variants at 189, 217, 247, and 261 as well as the tiny deletion. The other, less frequent but obviously related sequence had variants at 189, 217, and 261 but not 247. As film after film peeled itself out of the developing machine over the next few weeks, I got very good at recognizing the particular pattern of bands that meant we had found the Polynesian sequences. There they were, spread back along the island trail to Polynesia. The further west we went and the closer we got to the Asian mainland, the rare the full sequence with 247 became, while a new type with just 189 and 217 began to appear, reaching its highest concentration among the Ami, Bunum, Atayal and Paiwan from Taiwan. The record of the whole amazing journey was there. I rang as many people as I could think of who might have new mitochondrial sequences from native Americans. I had to be sure that 247, the defining variant of Polynesian mitochondrial DNA, was not abundant in the Americas. No-one had seen it. Not even once. Heyerdahl was wrong.
Why there was a "long pause" before ancient humans completed their colonization of islands in the Pacific Ocean

WIND BLOWN (BLOWN OUT)

I could not help feeling a tinge of disappointment that I had been unable to vindicate the man who had inspired a generation with his voyage in Kon-Tiki. But there it was. His theory had wilted under the fierce spotlight of genetics. The major opinion had been proved right: the Polynesians had come from Asia and not America. I never got to know what Heyerdahl himself thought about this. I am sure that, at eighty-three, he has better things to do than defend himself against the awesome power of modern genetics. There was a ripple of applause from the anthropology establishment when we published our results; but these academics were already so sure of themselves and convinced by the weight of the evidence for an Asian origin that they were not noticeably excited by this new information. To agree with the prevailing consensus is unlikely to disturb the peace. To disagree with it, as I was to find out before long in another part of the world altogether, was anything but peaceful.
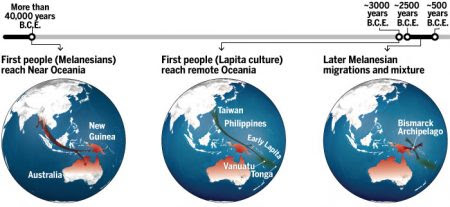
The genetic trail into the scattered islands of the vast Pacific was now crystal clear. The ancestors of the Polynesians began their epic journey in either coastal China or Taiwan. This is where the highest frequencies of what we can safely assume to be the ancestral mitochondrial DNA sequence of most Polynesians are found today, with variants at 189 and 217 and the small deletion. We also found in the samples from Taiwan other sequences with extra variants on top of the core 189, 217 pattern but at positions we didn't find in other parts of the region. These are the mutations that have happened in Taiwan since the ancestors of the Polynesians left. By counting up the mutations and multiplying by the mutation rate we can estimate the length of time since the ancestral sequence itself arrived in Taiwan. As we shall see when we come to explore the genetic landscape in Europe, this is a controversial area in conemporary research. None the less, it was pretty clear from the great diversity of variation on the basic theme of 189, 217 in Taiwan that the sequences had been there a very long time indeed, probably as long as twenty to thirty thousand years.
There are many archaeological signals of a very sudden population expansion in the islands of south-east Asia around three to four thousand years ago, defined by a range of artefacts associated with an agricultural economy. The most significant of these is pottery of a distinctive style called Lapita, with a red surface glaze and tooth-like decorations stamped into the clay in horizontal lines. For archaeologists, pottery with an identifiable style is a real bonus. It survives for thousands of years in the ground, and a similarity of ceramic style can connect settlements that are geographically far apart. It doesn't automatically mean that the peopl who used the pottery were biologically related, but it is a certain sign of contact between the different places. Within a period of only five hundred years, beginning three and a half thousand years ago, Lapita settlements appeared on the coast of many of the islands in the western Pacific, from the Admiralty Islands north of New Guinea to Samoa in western Polynesia. Supporters of the Asian origins of Polynesians had always associated this rapid expansion, which implied a sophisticated seagoing capacity, with the people who ultimately colonized the whole of Polynesia. The mysterious absence of Lapita pottery on the islands to the east of Samoa was explained by the lack of a suitable clay. Now that the genetics has come down firmly in favour of an Asian origin rather than an American origin for the Polynesians, could we say anything new about where this remarkable expansion of people and pottery might have begun?
First of all, the complete absence of the variant at position 247 in Taiwan made it extremely unlikely that it had started there. If it had, then I would have seen plenty of variant 247 in Taiwan. In fact, I never see variant 247 west of Borneo. So the rapid Lapita expansion is only supported by the genetics if it began somewhere to the east of Borneo. I did see 247 inn the Moluccas, an island group in Indonesia, and it has been there long enough to accumulate additional mutations. My best estimate for the place of origin of the remarkable Lapita Polyneisians would be somewhere in that island group. From there, the mitchondrial trail leads out into the Pacific, to Hawaii in the north, to Rapanui (Easter Island) in the east and to Aotearoa (New Zealand) in the far south.
All this is clear from the main Polynesian type. But what of that strange, rare sequence that I had found in the blood from a single outpatient in Avarua hospital and Koji Lum had found in one native Hawaiian? Could this be the faint echo of Heyerdahl's American Polynesians? We had certainly found this sequence all over Polynesia after our extensive sampling, though it was never common; but none of my contacts had seen anything like it in North or South America. Then we found a single example in Vanuatu and two more from the north coast of Papua New Guinea. However, only when I tracked down some old samples from the mountainous interior of New Guinea did I find this sequence in abundance. This was mitochondrial DNA that had been handed down to the present-day inhabitants from the earliest settlers of that huge island - settlers who, according to the dating of early archaeological sites, had made their way there at least forty thousand years ago in the same ancient migration that had carried the first Australians to that vast continent. So the direct maternal ancestors of the mysterious outpatient from Avarua hospital had spent almost forty thousand years on the island of New Guinea before joining a Lapita voyaging canoe heading east into the unknown.
From the north coast of New Guinea a line of islands, each visible from the previous one, stretches out into the Pacific as far as the Solomon Islands. These are high islands with mountain peaks which can be seen on the horizon either before setting out or, at the very least, before losing sight of your departure point. This comparatively safe navigational technique had already taken the earliest settlers of New Guinea up past New Britain and New Ireland and down the main chain of the Solomon Islands as far as San Cristobal thirty thousand years ago. But this was the end of the pier. Beyond that was the end of the sea with the nearest land, the islands of Santa Cruz, still three hundred kilometres away far beyond the horizon. There is no archaeological evidence of any settlement beyond the Solomons until the arrival of the Lapita people twenty-seven thousand years later.
Two crucial developments enabled the new wave of colonists to launch into the unknown. The first was the development of the double-hulled voyaging canoe. These magnificent vessels reached enormous size. The first Europeans to reach Polynesia saw canoes over 30 metres long, and smaller versions are still used today. The double hull prevents capsizing in the same way as the outrigger on a catamaran. The vessels had a prow at each end, and so could be tacked across the wind and then reversed without turning around. These were the vessels that carried the Polynesians into the Pacific; the complementary and equally crucial development was a highly sophisticated set of navigational skills. Whereas the earliest settlers had managed to reach Australia, New Guinea and the Solomon Islands by steering to visible targets, the Polynesians sailed off into a void, not only unable to see land but not knowing if there was any. Their progress can be followed through the dating of archaeological sites. They quite quickly settled Santa Cruz and the islands of Vanuatu, paused before the 750 kilometre crossing to Fiji and beyond to Samoa and Tonga, then paused again before pushing on to the limits of Polynesia. They reached Easter Island and Hawaii about fifteen hundred years ago, and, last of all, New Zealand about twelve hundred years ago. They had reached every island in this vast ocean in a little over two thousand years. How did they do it?
Well supplied with food and water, the canoes set off against the prevailing wind. This might seem like hard work, but it did at least ensure a relatively safe return journey, sailing back downwind to the home port, navigating by the stars. It is fairly straightforward to travel along a line of latitude by keeping a rising or setting star in the same position relative to the canoe each night. When it is time to return home, it is a simple matter of reversing direction and sailing downwind guided by the same stars - simple in theory, but in practice still fraught with danger. It must have been all too easy to miss the home island, especially if it was passed in the night or in a storm. There must have been many losses.
More remarkable still are the signs the Polynesians used to detect the presence of unseen land. Cloud formations above high islands betray their presence over the horizon. The opalescent blue-green of low-lying atolls is reflected in the clouds under the right conditions. The outward and homeward flight directions of birds known to nest on land give clues. Floating debris shows there is land upwind. These are the visible signs. But early voyagers not only saw their way ahead; they felt it too. Even now traditional navigators can detect the change in the way the sea moves. The underlying swell sweeps across the ocean, but if reflected back from the islands- just as ripples from a stone tossed into a pond bounce back from the edge. Even a hundred miles from land an experienced navigator will use his feet to feel for the interference pattern as one swell crosses another.

A new genetic study shows some Polynesians share DNA with an ancient indigenous group in Colombia. This is fascinating and intriguing!
Also appears to be another blow to poor Christopher Columbus and his special distinction for "discovering" places people already lived
That is how the Polynesians arrived. It is not why. What drove them to these quite extraordinary feats of exploration is still a mytsery. It cannot be that they needed to keep moving on to satisfy the economic needs of a growing population. Many of the islands en route to the remote Pacific are large and fertile, and are not densely populated even today. Perhaps it was an insatiable urge to explore the unknown. They must certainly have voyaged right across the Pacific to reach South America. We know this from the evidence of the Kumara, the sweet potato, which was and is cultivated all over Polynesia. There is no dispute that the sweet potato is a true Andean crop, so this has to mean there must have been at least some contact between the native South Americans and the Polynesians. The genetics rules out Thor Heyerdahl's explanantion that the sweet potato was brought into the Pacific by the people who he thought had colonized Polynesia from South America: we had proved beyond any doubt that the colonization of the Pacific had happened in the opposite direction. For the sweet potato to have been imported from South America, the Polynesians must have found their way right across the Pacific. But they left no visible genetic trace in South America. To my knowledge, not a single sample of Polynesian mitochondrial DNA has ever been found there. However, I did eventually find two mitochondrial sequences from Tahiti in French Polynesia that matched published sequences from Chile. I like to think these may have been the faint echo of women who had joined the Polynesians on their return journey back into the Pacific from South America, but I cannot prove it.
.jpg)
Travelling so far to the south, the voyagers would have left themselves little hope of return had they not discovered land. Getting to Aotearoa was not a simple matter of sailing into the wind along a line of latitude in the knowledge that if no land was encountered it was only necessary to reverse direction and travel home along the same line with the wind behind them. The voyage to Aotearoa took them across latitudes and far to the south of the reliable trade winds into a part of the ocean where the winds were far less predictable. This was another level of maritime exploration altogether, of such maturity and daring that I feel sure the Polynesians must also have reached the coast of Australia, so accomplished were they in the ways of the ocean. But if they did land here, they left no trace. Perhaps they only felt comfortable settling uninhabited lands. Did they, I wonder, sail south of Australia and right across the Indian Ocean to Madagascar - unpopulated then, and now at least partially inhabited by people speaking a similar language? Could they have done that? I'm quite sure they could. Did they? One day, the genes will tell us.
I still feel a sense of excitement when I think about the work in Polynesia. I had found myself on an islands in the middle of the Pacific staring out at the ocean beyond the reef and overcome by a burning desire to find the answer to a question. I really wanted to know where the Polynesians had come from. It was a quest born out of pure curiosity; and it had delivered the answer, clear-cut and unequivocal, in a little over three years. Having seen how decisively mitochondrial DNA had settled the matter of Polynesian origins, I was very confident of its inherent ability to solve even harder questions in an arena much nearer home.
The Seven Daughter of Eve: The Science that Reveals our Genetic Legacy. Sykes, p. 96-107

Savings boost survival:food/resources get you through lean periods.Prudent to provide buffer against storm & starvation.Do all realise that?
http://www.youtube.com/watch?v=A69Z8fCpuAI
Hey Polynesians, Many Of You Live In Cold Climates Where You Wake Up Early And Work In An Office-Like Setting For Hours On End, But You Have Adaptations (Polynesian Genes) That Your Ancestors Evolved Specifically For Hunting And Gathering In The Humid Atmosphere Of Polynesia. One Of These Adaptations (Polynesian Genes) Is Responsible For Your Lazy, Procrastination Prone Disposition.
Why Expend A Lot Of Energy Planning, Building, And Maintaining Shelter When The Temperature In The Environment You Live Isn't Unbearable And Doesn't Fluctuate Wildly, And You're Constantly On The Move And The Shelter You Do Build Often Succumbs To The Elements? Wouldn't It Be Wiser To Build Less Elaborate, Temporary Shelters And Then Spend The Rest Of The Time Searching For Food, Searching For Mates, Sleeping, And Conserving Energy (Being Lazy*)?
Why Waste Your Energy (Bodily Resources) For No Reason? If You're Not Searching For Food Or Searching For Mates Or Avoiding Predators Or Attacking Enemies, Why Waste Your Energy? You Shouldn't And We Didn't In Our Evolutionary Past Because Doing So Would Put Our Lives In Jeopardy (If You're Wasting Your Energy On Meaninglessness And Food Is Hard To Come By You're Putting Yourself At Risk Of Dying From Starvation). So We Evolved To Be Lazy. It's Adaptive To Lie Around And Not Expend A Lot Of Energy.
*The Genes That Code For Sloth And Indolence Were Selected For In Ancestral Polynesian Populations And Are Now At A High Frequency Within These Populations. That Is, Polynesians Are Innately Lazy To A Greater Degree Than Other Races.

BONE BONE BONE BONE BONE...BONE BONE...BONE!


Hardwired for laziness? Tests show the human brain must work hard to avoid sloth @ubcnews #Neuropsychologia @ELSneuroscience

https://www.washingtonpost.com/news/in-theory/wp/2016/08/24/in-defense-of-leisure/?utm_term=.25663beef54d&wpisrc=nl_theory&wpmm=1
"You Slow And Lazy And You Kant Fade Me! That's The Reason I Knocced YO Ladie!" - Pimpsy In Reference To Pimp C!
THERE'S A PASSAGE IN THIS BOOK http://www.amazon.com/Mean-Genes-Taming-Primal-Instincts/dp/0465031242/ THAT WILL CORROBORATE WHAT I'VE WRITTEN IN THE POST ABOVE. I'LL COPY THAT PASSAGE LATER!
Most animals are lazy. Take mice, for example. Scientists investigating the effects of extreme exercise naturally want mice that are long-distance runners. The trouble is, mice don't want to run marathons any more than most humans.
When mice are placed on tiny treadmills, many of them will simply go on strike. They will even sit on the moving belt to the point that the skin on their butts begins to get scratched and scraped. The mice are ingenious in their ability to avoid exercise, positioning themselves against walls, splaying their feet at extreme angles - anything to avoid jogging. Any one of us who has made elaborate excuses to escape a workout can empathize.
In addition to orangutans like Chantek, the Yerkes Center has a large number of chimpanzees. Predictably, this population of zoo primates is also overweight and lazy. One of the matriarchs is a female called Natasha, known because of her bulk as Na "tank"-a.
Even though the chimps are very well fed, most of them still excitedly yell and run around if a person shows up with a box of oranges. Natanka is not so easily moved. She plants herself directly below the fruit platform and begs with subtle arm movements that require no more energy than changing the TV channel with a remote control. She will move only about six inches to snatch a ripe fruit; getting her to travel across the compound requires dozens of oranges, each artfully thrown a few inches ahead of her.
Laziness is good for most animals...To understand this, we have to leave our couches and think like wild primates. Energy in the form of food is hard to obtain, and once gotten, not to be squandered.
This why lions sleep most of the day, mice squat on treadmills, and people avoid the gym like the plague. The only people who exercise in poor countries are the privileged - the rich, the tourist, or the full-time athlete. Similarly, modern foragers lead active lives, but the concept of unnecessary exercise puzzles them.
Evolution favors the frugal and casts a hard, wary glance on an organism that frivolously wastes energy. What happens to animals that expend energy needlessly? They die, and their genes die with them. We descended from humans who were frugal with their physical activity, and we carry their energy-conserving genes.
Nature abhors wasteful energy expenditure but will induce us to work under many conditions...Those frequently sleepy lions will sprint if there is a gazelle to be chased or a hyena to attack. If we can set up situations with appropriate rewards, we can similarly shed the cloak of laziness...(Mean Genes)
This Is ME Every Night, But Rather Than Having A Laptop On My Gut And A Remote In My Hand I Have A Beer In Hand, Book On My Gut, And A Radio Nearby (Listening To Talk Radio).
Anthropologists have long debated how much work people had to do to achieve subsistence in preindustrial societies. The anthropological tradition prior to the 1960s assumed that hunter-gatherers lead hard lives of constant struggle to eke out a living. The Neolithic agricultural revolution, by increasing labor productivity in food production, reduced the time needed to attain subsistence, thus allowing for leisure, craft production, religious ceremonies, and other cultural expressions.
However, systematic time allocation studies of hunter-gatherer and subsistence cultivation groups undertaken from the 1960s onward revealed labor inputs in these societies to be surprisingly small. For example, the Hiwi, a foraging group from Venezuela, consumed a modest 1,705 kilocalories per day and often complained of hunger. Yet men would generally forage for less than 2 hours per day, even with high returns from each hour of work.
Indeed work time in these societies is considerably less than that in settled agrarian societies. Table 3.12 also shows estimates of the total work input of males per day in modern societies in which foraging and hunting were still significant activities. For these societies median hours of work per day for males, including food preparation and child care, were just 5.9 or 2,150 hours per year. Thus males in these subsistence societies consume 1,000 hours more leisure per year than in affluent modern Europe.
Such low work inputs need not be maladaptive for foragers. Ecologists have calculated for how many hours a day various bird and mammal species engage in "work" - foraging, moving, defending territory, or even socializing - as opposed to resting. If we take just the species closest to man - apes and monkeys - work hours per day averaged only 4.4.
The typical low work effort of subsistence societies helps explain why Polynesia appeared such an idyll to European sailors, and why captain Blyth had trouble getting his sailors on board again after their stay in Tahiti. The main food supplies in Polynesia were from breadfruit trees and coconut palms, supplemented by pig meat and fish. But all the labor that was required for the breadfruit trees and the palms was to plant the tree, tend it until it grew to sufficient height, and then harvest the fruits when ripe. Like the subsistence societies of table 3.12 the Polynesians apparently labored little.
A Farewell to Alms: A Brief Economic History of the World. Clark, p. 65-66.
Harpending...In a talk he gave at a 2009 conference on "Preserving Western Civilization," he asserted that people of sub-Saharan African ancestry have no propensity to work when they don't have to - "I've never seen anyone with a hobby in Africa," he said - because, he thought, sub-Saharan Africans have not gone through the type of natural selection for hard work in the last thousand years that some Eurasians had.

https://www.youtube.com/watch?v=ZRDaPEaDJ7E
The Protestant Work Ethic. White Americans Who Trace Their Ancestry To Western Europe Descended From Populations That Practiced An Agrarian Form Of Subsistence (Subsistence Farming). The Culture And Lifestyle Associated With This Form Of Subsistence (Careful Planning And Arduous Work For Extended Periods Of Time) Created Selection Pressures For A Certain Temperament And Behavioral Traits...TO BE CONTINUED
TABLES 3.11 AND 3.12
https://lesacreduprintemps19.files.wordpress.com/2012/12/the-10000-year-explosion-how-civilization-accelerated-human-evolution-2009-by-gregory-cochran-henry-harpending.pdf
WHY WHITEY WORKS. PAGE 110.
(Mane Wit Ho)

WHITEY'S BEEN WORKING HIS WHITE ASS OFF FOR FAR LONGER THAN THE INCEPTION OF THE INDUSTRIAL REVOLUTION, BUT SINCE THE INDUSTRIAL REVOLUTION, WHITEY'S WORKLOAD AND HENCE WORKING HOURS HAVE INCREASED STEADILY. CONSEQUENTLY, THE GENES THAT LED TO ECONOMIC SUCCESS IN THIS HEAVY WORKLOAD ENVIRONMENT (i.e. GENES THAT LEAD WHITEY TO HAVE A STRONG WORK ETHIC (THE PROTESTANT WORK ETHIC) AND BE INDUSTRIOUS) SPREAD EXPONENTIALLY THROUGHOUT THE WESTERN EUROPEAN POPULATION. SO, IN THIS CURRENT, MODERN ENVIRONMENT, THAT WHITEY CREATED, WHITEY IS MUCH MORE ENTHUSIASTIC ABOUT WORKING, EAGER AND EXCITED TO WORK, AND AT EASE* WITH WORKING LONG HOURS.
*AT EASE, FEEL THE BREEZE...SIPPIN' ON GINSENG AND JUICE!
*AT EASE, FEEL THE BREEZE...SIPPIN' ON GINSENG AND JUICE!

HUNTERS, GATHERERS, PASTORALISTS, HORTICULTURALISTS, AND CERTAIN AGRICULTURALISTS (i.e. THE ANCESTORS OF ALL BLACK AND BROWN PEOPLE THAT ARE ALIVE TODAY) DEVOTED LESS TIME TO ACQUIRING AND CULTIVATING FOOD AND MORE TIME TO REST AND RELAXATION. SO THE GENES FOR IDLENESS, INDOLENCE, AND PROCRASTINATION ARE MUCH MORE COMMON AMONG THEM (MODERN DAY BLACKS AND BROWNS) THAN THEY ARE AMONG WHITES, PARTICULARLY THOSE WHITES THAT TRACE THEIR ANCESTRY TO WESTERN EUROPE.
https://www.youtube.com/watch?v=A69Z8fCpuAI Jared Taylor
Jared Taylor
..@genophilia @racehist @ramzpaul
We are aesthetically unique and we created the civilization every other race covets.
Football, Like All Sports And Other Forms Of Entertainment, Doesn't Impact Humanity, Society, And The World As A Whole In An Overwhelmingly Positive Way, At Least Not As Overwhelmingly Positive As Science (Academia)! And Why Are There More Polynesians In The Sports World (Athletic Industry) Than The Scientific World (Academia)? Because Of The Natural Selection And Sexual Selection Forces Operating On Them Throughout Their Evolutionary Past. In Other Words, High Athletic Ability Was Of Greater Necessity In Their Environment Of Evolutionary Adaptedness (Polynesia) Than Was High Intelligence. So The Polynesians Of Today Are Stuck Coping With The World With Their Low IQs (The Demand For High Athletic Ability Was More Imperative Than The Demand For High Intelligence To Survive And Reproduce In Ancestral Polynesia).
You're EXACTLY Right, Kaba. Your Genes Account For About 80% Of Your Athletic Ability Just As They Account For About 80% Of Your Mental Ability (Intelligence). Your Environment (Uprbringing, Household, Schooling, Training, Etc.) Is Only Responsible For About 20% Of Your Athleticism And Intelligence In Adulthood. So All Of The Training In The World Won't Make You A Superior Athlete And All Of The Schooling In The World Won't Make You A Superior Intellectual Unless You Have The Superior Athletic Genes Or Superior Intelligence Genes To Accompany That Training And Schooling. (A Father Will Often Wonder "How Come He's Not As Good Of An Athlete As He Should Be? We Practice All Of The Time!" He's Not As Good Of An Athlete Because You Didn't Breed With A Female That Has Genes For Superior Athleticism, Dad. So You're To Blame, Dad. All Of The Training In The World Won't Make Up For His Genetic, Athletic Inferiority!
 |
| Sum Moe |
Why Are HOMOSEXUALS, GAYS, FAGS, SAME SEX LOVERS, Etc. So Common Among Filipinos And Polynesian Ethnic Groups? It May Have To Do With The Sex Ratios Within These Ethnic Groups (Polynesians And Filipinos* Tend To Have Families With Disproportionate Numbers Of Males. They Tend To Come From Male Dominated Cultures). http://julianodea.blogspot.com/2006/07/is-homosexuality-due-to-genetic.html DAME UN BESO, MIJO! (I Have An Excerpt From Matt Ridley's Genome: The Autobiography of a Species in 23 Chapters Concerning The Possible Biological Causes Of Homosexuality That I'll Post Later.)
*Although Males May Outnumber Females Within Filipino Households And The Ethnic Group As A Whole, Filipino Females Hold More Sway And Power Over Filipino Males. This Is Most Evident In Their Relationships (Filipinas Wear The Pants In Boyfriend/Girlfriend Relationships And Marriages). The FILIPINA (PILIPINA) Is The Dominant Figure In Filipino Society. No Wonder They've Had A Female President And Other Female Political Leaders.
http://www.scienceagogo.com/news/20100104222002data_trunc_sys.shtml
http://psy2.ucsd.edu/~mgorman/Vasey2009.pdf
Speaking Of The Philippines, Why Is It So Corrupt?
The Same Reason Africa, For The Most Part, Remains An Undeveloped, Non-Industrialized, Non-Urbanized Country Applies To The Philippines. They Have All The Resources In The World, But Because Of Their, On Average, Low Intelligence In Relation To 1st World Countries, They've Been Unable To Convert The Majority Of These Resources Into Wealth, Status, And Power On A Global Level And Everything That Comes With Wealth, Status, And Power (Including A Better Quality Of Life And Better Life Outcomes For Its Citizens).
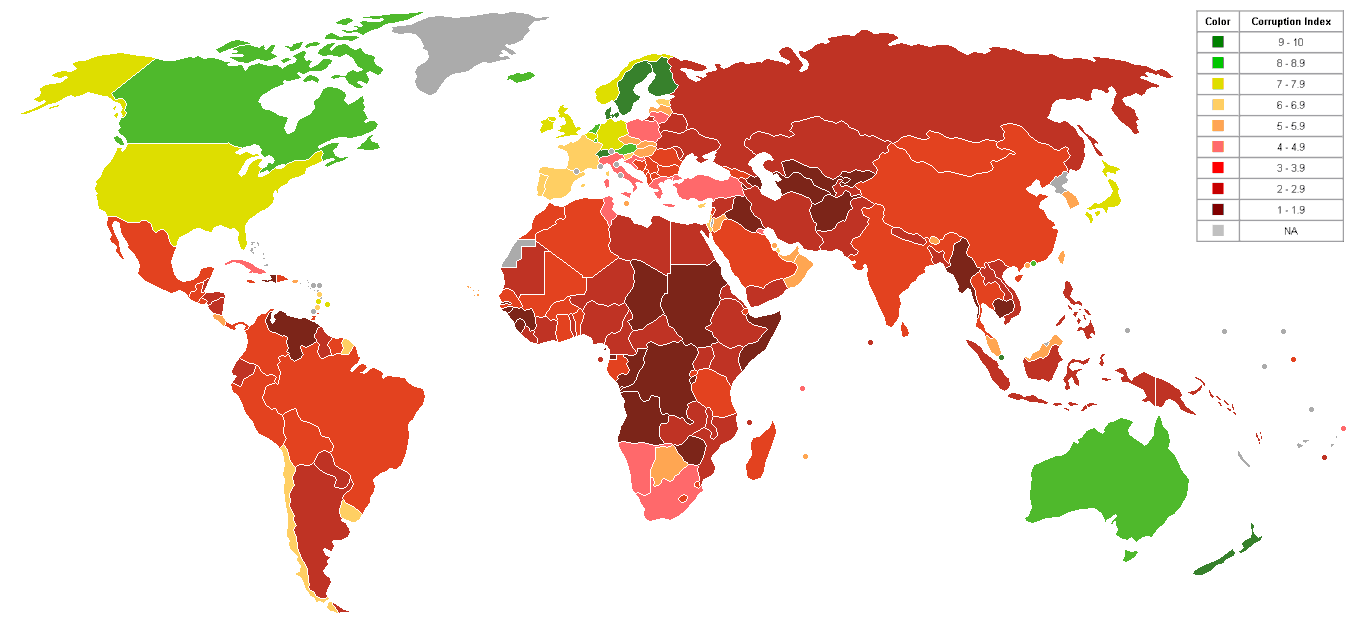
One's Level Of Intelligence Is Correlated With Personality Traits (Openness, Agreeableness, Conscientiousness, Extraversion, Neuroticism) That Determine A Group's Level Of Religiosity And Morality. Since Filipinos Have A Low Average Population I.Q. They Tend To Be More Religious, Yet, Ironically, Less Morally Inclined (As Exemplified In Their High Levels Of Political And Financial Corruption). Both Of Which (High Religiosity And Low Morality) Work In Tandem To Stifle Economic, Technological, Social, Etc. Growth. http://upload.wikimedia.org/wikipedia/commons/0/04/World_Map_Index_of_perception_of_corruption.png (Look At How Corrupt The Philippines Is.)
{kind=link}
http://www.sq.4mg.com/corrupt.htm
The Philippines Is The 37th Most Corrupt Country In The World!
http://www.psychologytoday.com/blog/the-scientific-fundamentalist/201005/iq-and-the-values-nations
Hey, Why Is The Philippines Such A Corrupt Country? Because Of The Filipino Population's Low IQ. This Is The Main Reason They Tend To Be Highly Religious, Yet Morally Backward And Sexually Promiscuous. (Highly Religious http://hypnosis.home.netcom.com/iq_vs_religiosity.htm Morally Backward. They're As Religious As They Are Intelligent.)